2013
Inicio

En un día muy soleado, un árbol de tamaño mediano (digamos: unos 10 m de alto y con un diámetro de tronco de 20 cm) puede transportar 200 litros de agua o más, desde el suelo hasta su copa, sin la intervención de mecanismo exterior de bombeo alguno. Entonces, ¿cómo es que la planta logra hacer circular el agua que cada parte suya requiere? El fenómeno de la capilaridad (cuadro 1, figura 1) explicaría una parte de esta función, pero no lo haría completamente, ya que dicho fenómeno sólo sería capaz de elevar el agua unos cuantos metros por arriba del suelo.

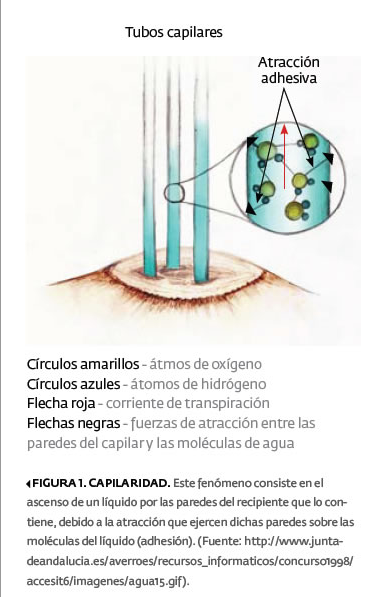
La teoría que mejor explica el ascenso del agua en las plantas, es la llamada coheso-tenso-adheso-transpiración, comúnmente reducida a coheso-tenso-transpiración o CT . A continuación, se presenta una descripción de esta teoría, a partir del análisis de sus componentes.

El fenómeno del ascenso del agua desde las raíces de una planta hasta sus hojas ha tratado de ser comprendido desde hace siglos. La primera persona que intentó explicar experimentalmente este fenómeno, con árboles vivos, fue Stephen Hales (1677-1761), clérigo inglés aficionado a la ciencia, quien se interesó en el estudio comparativo de la circulación de la sangre en los animales y del agua en las plantas. Además de ser el primero en medir la presión arterial en un animal vivo, estableció la naturaleza del movimiento del agua en las plantas, al descartar la presencia de un sistema de bombeo en ellas (figura 2).

Aunque Stephen Hales,2 a principios del siglo XVI, propuso que el agua absorbida por las raíces pasaba a través de los vasos, no tenía claro el proceso de transpiración de éstas. Cerca de finales del siglo XIX, los investigadores alemanes Boehm, Sachs y Strasburger concluyeron que la pérdida de agua a través de las hojas era el impulso necesario para que se produjera el ascenso del agua. Finalmente, en 1895, Dixon y Joly 3 propusieron que el agua dentro de los vasos tiene una gran fuerza de cohesión y puede estar sujeta a grandes tensiones.
Los vasos forman parte del tejido vascular de las angiospermas (plantas con flores), por los cuales circula el agua, así como algunos minerales y compuestos orgánicos disueltos en ella (la savia bruta). El otro componente de este tejido es el floema, por donde circula la savia elaborada (azúcares, principalmente). Los tejidos vasculares conectan las raíces con el tronco y a éste con las ramas de grande y pequeño calibre, las cuales, a su vez son vinculadas con las hojas.
Coheso-tenso-transpiración

Las plantas consumen grandes cantidades de agua; la mayor parte de ellas pasa breve tiempo en los tejidos vegetales, antes de emitirse a la atmósfera. El agua que queda en las hojas se usa en la fotosíntesis, para construir nuevos tejidos.
Esta teoría (CT) debe su nombre a las fuerzas a las que se encuentra sometida la savia dentro de los conductos de las plantas: 1) La fuerza de cohesión que existe entre las moléculas de agua, la cual es resultado de la atracción de dos cargas opuestas —la del H y la del O—; 2) la transpiración de las hojas, que jala la savia hacia arriba; 3) desde abajo, la fuerza de atracción de la gravedad jala la savia hacia el suelo, generando así una fuerza de tensión.
Las paredes de los vasos no son lisas, sino que presentan relieves más o menos complejos —dependiendo de la especie—, los cuales aumentan la superficie interna de los vasos, donde las moléculas de agua se adhieren, resistiendo así la fuerza de gravedad.
Esta combinación de fuerzas: la de cohesión entre moléculas de agua y la adhesión de ellas a las paredes de los vasos conductores permite que se formen largas y continuas cadenas de agua desde las raíces hasta las hojas. El agua que se evapora de la superficie de las hojas es remplazada por el agua almacenada en su interior, que a su vez es sustituida por el agua contenida en las ramas. El agua que se pierde en el tronco se suple con la de las raíces, pues éstas se encuentran absorbiendo constantemente agua del suelo.
El potencial hídrico

El potencial químico es una medida del estado de energía de una sustancia (es decir, de su capacidad para realizar un trabajo). En el caso del agua, se conoce como potencial hídrico, y se representa por la letra griega Psi (ψ) acompañada del subíndice “w” (por water: agua, en inglés). El ψw del agua pura —sin sales— es cero. Al añadirse solutos al agua, su potencial hídrico se va haciendo más negativo. Algunos valores típicos de potencial hídrico se presentan en la tabla 1.
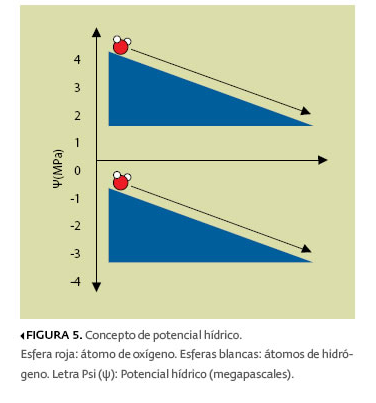

En el interior de la planta, el agua se mueve siempre en dirección de mayor a menor valor de potencial hídrico; es decir, de más negativo a menos negativo (figura 5), y la diferencia absoluta de los valores de ψw constituye un gradiente —o sea, un cambio que ocurre gradualmente—. Como se aprecia en la tabla 1, los valores de ψw que encontramos tanto en el suelo como en la planta y en la atmósfera son negativos. La diferencia entre el ψw del suelo y el de la atmósfera puede ser de dos a tres órdenes de magnitud.
El reducido ψw del suelo es ocasionado por la presencia de sales en el agua que ahí se encuentran y por la atracción que ejercen las partículas sólidas del suelo sobre las moléculas de agua,4 mientras que el gran ψw de la atmósfera se debe a que, en ella, las moléculas de agua se encuentran muy alejadas una de otra (estado de vapor), reduciendo así las fuerzas de atracción intermoleculares. Por lo tanto, el ascenso del agua en la planta ocurre desde la solución acuosa del suelo (valores negativos menores) hasta la atmósfera (valores negativos mayores), pasando a través de la planta (valores intermedios).
Eficiencia y seguridad en la conducción del agua

No existe una velocidad uniforme en el proceso de conducción de agua en las plantas; una misma planta experimenta diferentes velocidades durante el día, dependiendo de las condiciones atmosféricas (temperatura, velocidad del viento, intensidad luminosa, etc.). Durante la época seca del año, las plantas pierden agua rápidamente por las hojas, mientras que en el suelo hay poca agua. Esto causa que las columnas de agua dentro de las plantas sean sometidas a mayor tensión; si ésta rebasa la resistencia de la columna, se puede romper, e interrumpir así el flujo de agua hacia las hojas. Este fenómeno se llama cavitación.
La resistencia que pueda tener una planta a la tensión de las cadenas de agua depende de la estructura de su sistema vascular. Generalmente, los vasos gruesos son muy eficientes conductores, pues transportan mayores volúmenes de agua que los más delgados. No obstante, también están sujetos a mayor tensión que los vasos delgados, pues el mayor volumen transferido de agua implica mayor peso. Este peso del agua puede ser mayor que la fuerza ejercida por la transpiración y que la adhesión del agua a las paredes de los vasos, causando que las cadenas de agua se rompan. Cuando esto sucede dentro de un vaso, se dice que está cavitado. En la naturaleza encontramos una combinación de vasos anchos y delgados en las plantas, lo cual se debe a que éstas se enfrentan constantemente a la fluctuación del equilibrio entre eficiencia y seguridad en el abasto y conducción de agua.
Vulnerabilidad del sistema hidráulico

Para romper la cohesión de una cadena formada por moléculas de agua, se necesita ejercer una tensión igual a -20 MPa (megapascal, unidad de presión) y, para entender la magnitud de esta tensión, imaginemos que llenamos una jeringa con agua pura, libre de partículas sólidas y de gases; si selláramos el lugar donde se conecta la aguja y del émbolo colgáramos un peso equivalente a dos veces el peso que ejercería una columna de agua del mismo diámetro que la jeringa, pero con una longitud mínima de 2 metros, lograríamos romper la columna de agua dentro de la jeringa; esto, en el caso ideal de que el interior de la jeringa estuviera libre de imperfecciones.
En realidad, la tensión necesaria para romper las cadenas de agua dentro de los vasos de un árbol es mucho menor que en el caso hipotético de la jeringa, pues por los vasos de las plantas no fluye agua pura, desgasificada; todo lo contrario: contiene disueltos minerales y algunas impurezas, además de gases; esta solución se llama savia bruta y, aquí la llamaremos simplemente savia.
Dentro de los vasos, puede ocurrir un fenómeno llamado nucleación, que es la incorporación súbita de una microburbuja de aire a la savia. Estas microburbujas de aire pueden provenir de microfracturas en las paredes celulares de los propios vasos o de las células vecinas. De cualquier manera, al entrar una de estas microburbujas al torrente de circulación de la planta —que se encuentra bajo tensión—, la burbuja se expande, desplazando el agua dentro del conducto. Si la burbuja es tan grande que llena todo un vaso, éste se llega a cavitar completamente, como se explicó antes. Al llenarse de aire todo el vaso, se interrumpe el flujo de agua y, si la mayoría de los vasos se llegan a embolizar —es decir, se paraliza en ellos el suministro de agua—, la planta puede marchitarse por completo.
Importancia de estudiar el transporte de agua

Las plantas, por tener un sistema de circulación abierto (el que permite al agua entrar por las raíces y, eventualmente, salir por los estomas de las hojas —a diferencia del sistema vascular de los animales, que es cerrado, pues en él la sangre circula y se recicla—), son susceptibles a la sequía de la atmósfera: entre más alta sea la temperatura ambiente, mayor será la pérdida de agua por las hojas, lo que provocará el aumento en la tensión de la savia y esto causará un incremento en la frecuencia de embolismos. Sólo los sistemas vasculares más seguros, con un predominio de vasos delgados, serán capaces de sobrevivir en un ambiente demasiado seco; sin embargo, Olson y Rosell afirman que la frecuencia predominante de vasos delgados en climas secos se debe a que las plantas tienden a ser más bajas que las que crecen en climas húmedos; el diámetro de los vasos estaría directamente relacionado al tamaño de las plantas, más que al clima.5
Prevención a futuro

La comprensión del transporte de agua en las plantas es muy importante para poder entender su distribución geográfica actual y futura.
La distribución actual de las especies vegetales es el resultado de su adaptación al medio; por ejemplo, aquellas que han conquistado los ambientes más secos tienen un sistema vascular adaptado para soportar grandes tensiones en las cadenas de agua, evitando los embolismos.
Con el cambio climático, se espera que las zonas áridas se extiendan considerablemente en el planeta; entonces, ¿cuáles son las especies capaces de adaptarse a esos cambios?; sin duda, las que cuenten con un sistema vascular más seguro.
¿Qué mecanismos permiten a las plantas desarrollar vasos seguros contra la cavitación? Aún no lo sabemos, pero se está avanzando mucho en ese sentido. Indudablemente, tanto el diámetro como la distribución de los vasos están determinados por el ambiente y por los genes.
Así pues, es necesario encontrar los genes responsables de estas características del sistema vascular, con lo cual podremos empezar a reproducir las especies con las que habremos de hacer frente al calentamiento global.
Agradecimientos
Se agradece el apoyo del Conacyt (registro No. 165806) que me permitió escribir este artículo durante mi estancia sabática en la Universidad de Sao Paulo, en Brasil. La Dra. M. Lascuráin brindó valiosas sugerencias al manuscrito original.
Referencias
1. P. S. Nobel (2005). Physicochemical and Environmental Plant Physiology, 3a edición. Elsevier Academic Press, N. Y.
2. S. Hales (1961). Vegetable Staticks; Or, An Account of Some Statical Experiments on the Sap in Vegetables (Primera publicación: 1727). Macdonald & Co., N. Y.
3. H. H. Dixon y J. Joly (1895). On the Ascent of Sap. Philosophical Transactions of the Royal Society of London. B. 186: 563-576.
4. M. B. Kirkham (2005). Principles of Soil and Plant Water Relations. Elsevier Academic Press, San Diego, Ca.
5. M. Olson y J. Rosell (2013). “Vessel Diameter– Stem Diameter Scaling across Woody Angiosperms and the Ecological Causes of Xylem Vessel Diameter Variation”. New Phytologist 197: 1204–1213.
Autor
Guillermo Ángeles Álvarez es Biólogo por la Facultad de Ciencias–UNAM, Maestro en Ciencias por la University of Wisconsin-Madison y Doctor en Ciencias por el College of Environmental Science and Forestry. SUNY-Syracuse, N. Y., USA. Actualmente, es Investigador Titular en el Instituto de Ecología A.C., y miembro del SNI (I). Sus líneas de investigación versan sobre la estructura funcional de plantas tropicales, la anatomía de plantas leñosas y la eco-fisiología de plantas parásitas.
Fuente: CONACYT








